 המועדפים של davidfbird
המועדפים של davidfbird

















תמונות/קולות

מה
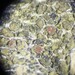
חיידקים (ממלכה Bacteria)מתצפת.ת
closterium_mysteriumתיאור
Video: https://youtu.be/YkORShmMZTk
Sampling location: The soil sample was collected from the bank of a pond on the Bagaryak River, near the water. The soil was wet but gradually dried out.
Date and time of collection: September 8, 2023
The sample was stored in a sealed glass container at room temperature.
On September 22, 2023, at 1 PM, a small amount of this soil was mixed with clean water.
Date and time of observation: November 22, 2023, at 7 PM
A number of clusters of bacteria in an unidentified particle of dead organic matter.
Rod-shaped bacteria that form star-shaped clusters. Each bacterium is constantly moving. So is each cluster as a whole.






תמונות/קולות

מתצפת.ת
rgeraldporterתיאור
F3 Hybrid Blue-winged Warbler x Golden-winged Warbler : as described in pp 156,158 https://www.westernfieldornithologists.org/archive/V42/WB-V42(3)-webComp.pdf
"This decision recognized evidence from the zone of hybridization that a black
postocular eyeline curling down around the auriculars is typical of F3 or F4 hybrids."



תמונות/קולות

מתצפת.ת
strangwayתיאור
Surface water grab from the Matchedash Bay - South Beaver Pond.

תמונות/קולות

מתצפת.ת
closterium_mysteriumתיאור
Observation of 14 specimens of Bicosoeca. This water sample was rich in this genus of flagellates. And in fact, there were even more, but I decided to limit myself to the most successful specimens from the microscopic point of view. I have no certainty that they all belong to the same species. If you can see that there is more than one species, please tell me about it and I will split the observation.
A water sample was taken from the bank of the Ufa River. The sample was stored at room temperature and observed 7 days after collection.
Video: https://youtu.be/Q2yi91jhIXE







תמונות/קולות

מתצפת.ת
peptolabתיאור
Spirostomum ambiguum Ehrenberg, 1834. from the superficial river edge benthos of the freshwater segment of the estuarine Peconic River. Imaged in Nomarsdki DIC on Olympus BH2S using Splanapo 20 0.70 and SPlanapo 40 0.95 objectives plus variable phone camera cropping on Samsung Galaxy S9+. The cell measures 960 um in length. The peristome is 2/3 the length of the body with the cytostome at the posterior 2/3 location. There is a large terminal contractive vacuole with a long collecting canal reaching almost the anterior end of the cell. The macronucleus is moniliform forming a chain of connected elongate spindle shaped macronuclear nodules beginning at the anterior 40% of the cell length and extending almost to the posterior end.
"Spirostomum ambiguum Ehrenberg, 1834. [syn: Trichoda ambiguum Müller, 1786; S. ambiguum var. major Roux, 1901] 900 m several mm long. Length: width ratio about 9-17. 15-25 kineties on each side; heterogeneous, numerous (4-5) CG rows per stripe. Peristome always longer than 1/2 of the body length, often reaching 2/3. CV much shorter than body length, rarely exceeding 1/10. The color depends on cytoplasmic granules. Moniliform MAC with 12-50 (avg. 15-25) nodules not exceeding 35 45 m in length when stained by Feulgen reaction. Numerous (up to 100) MICs 1-2 m long. Mono phyletic. Only found in freshwater. Reported in central and northern Europe, England, Russia, central Africa, USA, Jamaica, India and Japan. It sometimes harbors prokaryotic symbionts in the MAC. Spirostomum ambiguum is a well-defined, easily recognizable morphospecies whose monophyly is also strongly supported by molecular sequences" (1).
"Differential diagnosis
1) Size in vivo 1-000-4000 x 48-100 um, mostly 1200-2000 um. Visible with the naked eye as a white thread. Very flexible and contractile, shortens to approximately 390-430 x200-220 um. Contracted cells cigar-shaped.
2) Shape slender to moderately broad worm-shaped, more or less parallel-sided, 10-17 times longer than wide. Front end rounded, rear end truncated. Slightly flattened laterally. Ventral side in area of the mouth entrance slightly bulging.
3) Macronucleus moniliform or rosary-shaped, consists of 10-50, mostly 15-20 ellipsoids, about 18-53 x 12-24 um large nodes that form a long band that approaches the dorsal side. The number of nodes correlates positively with the age of the cell (REPAK & ISQUITH L974).
4) Contractile vacuole at the posterior end, with a long collecting duct extending forward along the dorsal side which sometimes shows ampullary extensions.
5) Close under the pellicle there are many spherical, spherical, yellowish granules arranged in elongated bands of 4-5 rows each which give the cell a yellowish to brownish color.
6) About 70-90 slightly spiraling rows of cilia, consisting of basal bodies arranged in pairs are constructed, but only the front one has a cilium.t
When the cells contract (startle reaction), the rows of eyelashes spiral around the body.
7) The adoral membrane zone extends from the anterior end to the posterior third (about 65-70 % of the body length) and turns to the right at the lower end. Parallel to the adoral membranelle zone a non-ciliated oral groove, which is bordered on the right by an undulating membrane.
8} Movement hatching, worm-like crawling and writhing. Fluidity with longitudinal rotation axis, with the front end describing a cone-shaped body of revolution. In the plankton falling floating with inclined longitudinal axis" (2).
"Spirostomum ambiguum is easy to recognize because of its size and shape, but the differentiation with S. minus causes considerable difficulties. There are many shape variations which occur, size and shape are not reliable distinguishing features. Usually S. minus is significantly slimmer and almost never reaches a length of 1 mm, while S. ambiguum is more compact, stockier and over 1 m long. Only the length of the adoral membrane zone in relation for body length (S. ambiguum: 65-70 %; S. minus: 35-50%) remains reasonably reliable distinguishing feature, but here too the differences do not seem to be too pronounced. Furthermore, the ciliates S. semivirescens have a similar size and shape (up to 2 mm, very slender, adoral membrane zone up to 50 % of body length, green through symbiontic algae) and Homalozoon vermiculare (up to 1.5 mm, 5-21 contractile vacuoles along the dorsal side, mouth small and only at the front end) as well as microturbellaria and nematodes. The characteristics 1,3 and 7 are particularly important for identification" (2).
- Focusing on Genera to Improve Species Identification: Revised Systematics of the Ciliate Spirostomum Vittorio Boscaroa,1, Daniela Carducci, Giovanna Barbieri, Marcus V.X. Senra, Ilaria Andreoli, Fabrizio Erra, Giulio Petroni, Franco Verni, and Sergei I. Fokin. Protist, Vol. 165, 527–541, August 2014
- FOISSNER W., BERGER H. & KOHMANN F. (1992): Taxonomische und ökologische Revision der Ciliaten des Saprobiensystems - Band II: Peritrichia, Heterotrichida, Odontostomatida. – Informationsberichte des Bayer. Landesamtes für Wasserwirtschaft, 5/92: pp 317-25.
תמונות/קולות

מתצפת.ת
mnold1תיאור
mag. 400x
- A pond-side water/periphyton sample was taken 04/22/2024 using a small sample bottled attached to an extension pole. Air temp 61F.
Long, juggling club-shaped cells, with siliceous scales, on stalks directed toward the colony center. Two flagella per cell, one long one short. This taxon has been previously observed in this general location, https://www.inaturalist.org/observations?place_id=any&taxon_id=494360&user_id=mnold1&verifiable=any.
Video shows a slowly rotating colony, https://youtu.be/nJn1INXd_co.
Ref. https://www.keweenawalgae.mtu.edu/gallery_pages/synurophyceans.htm



תמונות/קולות

מתצפת.ת
mnold1תיאור
Mag. 400x (1,4,6); 200x (2,3,5)
- A pond-side water/periphyton sample was taken 04/22/2024 using a small sample bottled attached to an extension pole. Air temp 61F.
Various forms of Hyalotheca dissiliens. The first 3 images show a mass of H. dissiliens zygospores. These masses are the largest I've encountered and were seen throughout the slide. The 4th image shows a segment of H. dissiliens with cells attached in filamentous form, surrounded by broad mucilaginous sheath. The last 2 images show H. dissiliens in single cell format. Both lateral views (barrel-shaped) and apical views (where the chloroplast has a stellate form) are visible.
Ref. https://www.outerhebridesalgae.uk/desmids/desmid-species.php?id=324


תמונות/קולות

מתצפת.ת
ikhomתיאור
A single egg in a poorly constructed nest on the ground. It was smaller, about 2/3 of a chicken egg.

תמונות/קולות

מתצפת.ת
peptolabתיאור
Paramecium nephridiatum V. Gelei, 1925 from the brackish estuarine pond Pussy's Pond, an offshoot of Acabonac Harbor. The population arose after feeding the culture with boiled wheat seeds. Imaged in Nomarski DIC on Olympus BH2S using SPlanapo 40 0.95 and SPlan 100 1.25 oil objectives plus variable phone camera cropping on Samsung Galaxy S9+.
The cells measure from 100-165 um in length. There is a large oval macronucleus usually situated slightly anterior to the cell equator and 3-4 small endosomal-type micronuclei can be observed with exact focus in the anterior end of the cell remote from the macronucleus- an important specific character of P. nephridiatum according to Fokin et al 1999 (1) The oral apparatus is situated posterior to the macronucleus with the cytostome roughly equatorial. There is an inconspicuous tuft of long caudal cilia. The two contractile vacuoles, one anterior and one posterior, have up to 15 short radiating canals and 2-3 excretory pores- another important specific characteristic of P. nephridiatum (1). There is a dense investment of subpellicuar extrusomes. The pellicle is divided into rectangular spaces on optical sectioning of the surface, a feature not mentioned by Fokin et al 1999. These features point strongly to P. nephridiatum. However, the more posterior location of the oral opening (as opposed to slightly anterior to the middle of the cell) and the peculiar pellicular division into rectangular spaces suggest that this might be a cryptic species of P. nephridiatum. Indeed, Przyboś et al 2019 describe 6 cryptic species within P. nephridiatum (2)
The cell shape with its rounded posterior end, and broad anterior part is of the "bursaria" type (Wichterman 1986) or "woodruffi" type according to Jankowski (1969). The oral opening is situated a little anterior to the middle of the cell. The cell surface is uniformly ciliated except for several long caudal cilia located close to the posterior end of the dorsal side, but not on the top. The abundant subpellicular trichocysts are distributed uniformly. Specimens were about 145 x 47 um long in vivo but shrank 10% after the silver nitrate impregnation. In silver-impregnated specimens, there were ~38 rows of ventral kineties and ~35 dorsal kineties. The preoral suture is distinct, but the postoral suture is very obscure. The cytoproct is situated in the posterior third. The vestibular zone is conspicuous and is terminated by the distinctive shape of the buccal opening. Two peniculi and open quadrulus are located on the dorsal and left wall of the buccal cavity. The endoral membrane is situated along the entire right edge of the buccal opening but its dikinetids are not recognizable in all specimens. It has 15 dikinetids on the average. The buccal cavity size various around 30 um. On the dorsal side of the body the contractile vacuole pores are very distinctive both on impregnated specimens as well as in living cells. Usually, both contractile vacuoles have more than one pore each, typically two or three. However, we have found two stocks (WCh-1 and WS-12) where one of the contractile vacuoles quite often (up to 50%) had only one pore. Both contractile vacuoles usually have 8-14 collecting canals, ten on average. Numerous crystals were very often found in the cytoplasm, but their quantity and location varied, probably depending on the culture conditions. The nuclear apparatus is located in the anterior part of the cell. One slightly ellipsoid or ovoid macronucleus, ~30x36 um in living cell and ~17x23 um in stained cells, on average, resides just anterior to the equator of the cell. In Feulgen-stained cells the macronucleus has a very intense colour. The three to four spherical micronuclei of the "endosomal" type (Fokin 1997), ~3 um in diam. (on average) are distributed irregularly along the anterior part of cell. Endocytobiotic bacteria are often found in the cytoplasm (Fokin 1989) and can also be found in the perinuclear space and in the macronucleus (Fokin 1989; Fokin and Sabaneyeva 1997)" (1).
"Swimming behavior - During swimming this species spirals on its long axis in both directions (Fokin 1987). We could not find any simple triggers (food, time) for changing this swimming direction. Typically, "left spiral swimmers" and "right spiral swimmers" were present at the same time in the culture. During the several years of investigation of this trait there was some preference of the cells from the same stocks to spiral in the left direction. Remarks- Gelei (1925) described a new species of Paramecium, Paramecium nephridiatum, based on the material which he had found in his laboratory aquarium. This population was in fact a mixture of the new species and Paramecium caudatum (Gelei 1938). This was the reason why some features of the new ciliate were similar to P. caudatum so that no one recognized this new species in nature, though reference to P. nephridiatum was made by Kahl (1931) and Kalmus (1931). Gelei (1938) redescribed the species from a native population (Tisza River, Szeged, Hungary) using a "clean culture". For unknown reasons, this new description did not attract the attention of protozoologists and in all subsequent reviews, P. nephridiatum was considered a nonvalid species (Vivier 1974; Wichterman 1953, 1986) even when the article of Gelei (1938) was listed in the references. Only once was P. nephridiatum mentioned in a short abstract as a species living in Florida, USA (Bovee 1983), although characters of the species were not listed in this publication. Since 1983, one of us (S. F) has repeatedly collected a species of Paramecium with multiple contactile vacuole pores, which is a distinctive trait of P. nephridiatum, although it was considered for a time as a feature of Paramecium woodruffi (Agamaliev 1983; Fokin 1986), Jankowski (pers. commun.)" (1).
"Occurrence and ecology- A number of stocks of P. nephridiatum were isolated from the sea shores of northern Europe: the North, Baltic, White, and Barents Sea coasts. It was detected during sampling in Woods Hole, MA, USA, Atlantic Ocean and on Sakhalin Island, Sea of Okhotsk. The salinity of these samples varied from 1.5-32 % The species was also found in a fresh-water body in Jerusalem Zoo, Israel. Samples were taken mainly during the summer, from mid-April (Wood Hole) to November (North Sea coast).
Sampling of the same wild population of P. nephridiatum (Sredny Island, White Sea, Russia) has been repeated every year since 1990 to observe long-time changes in the population, as well as the euryhaline ability of the species. This population was present at salinities from 4-35% and in the temperature range from 10-25” C. Very often the populations of P. nephridiatum occurred at the lower limit of oxygen concentrations. They were mainly feeding on bacteria. In the same samples these other ciliates were usually found: Prorodon sp., Frontonia marina, Metopus sp., P. calkinsi, P. woodruffi, and sometimes P. duboscqui" (1).
- Sergei I. Fokin, Thorsten Stoeck, and Helmut J. Schmidt; Rediscovery of Paramecium nephridiatum Gelei, 1925 and its Characteristics. J.Eukaryot.Microbiol. 46(4):416-426, 1999
- Evaluation of the molecular variability and characteristics of Paramecium polycaryum and Paramecium nephridiatum, within subgenus Cypriostomum (Ciliophora, Protista). Ewa Przyboś, Maria Rautian, Alexandra Beliavskaia, Sebastian Tarcz . Mol Phylogenet Evol . 2019 Mar:132:296-306


תמונות/קולות

מה
סוג Ulmusמתצפת.ת
emilitaתיאור
Covered in butterflies and flies. Didn’t seem to be leaking sap so I don’t know why.








תמונות/קולות

מתצפת.ת
closterium_mysteriumתיאור
Sampling location: A water sample was collected from Istok Lake near the shore, where snags and aquatic vegetation were present.
Several specimens of a freshwater sponge growing on different parts of Fontinalis antipyretica (water moss).
Date and time of collection: 16 Jul 2023 at 3 PM
Date and time of observation: 21 Jul 2023 at 7 PM
The sample was stored at room temperature in a plastic container.
During the initial days, the sponges were actively filtering, causing visible ripples in the water. However, by the day of observation, the noticeable water movement had ceased.


תמונות/קולות

מתצפת.ת
ikhomתיאור
On attached corticated Populus branch tiny black cups.
Next to Resupinatus applicatus.
Asci with many spores, IKI+.
Ascospores grey to brown with 1-septum, measured
*(5.1) 5.8 - 8.3 (8.7) × (2.4) 2.7 - 3.7 (3.9) µm
Q = (1.5) 1.8 - 2.6 (2.8) ; N = 17
Me = 7.1 × 3.3 µm ; Qe = 2.2




תמונות/קולות

מתצפת.ת
ikhomתיאור
On decorticated Eastern White Cedar - Thuja occidentalis branch, tiny cup fungi.
Apothecia sessile, with dark (black) margin and pale hymenium.
Asci 8-spored, IKI+ blue.
Ascospores hyaline, cylindrical, C-shaped, with 0-1 septum, measured
*(28) 29.4 - 33.5 (34.4) × (2.1) 2.5 - 3.3 (3.4) µm
Q = (8.1) 9.8 - 13 (14.7) ; N = 15
Me = 31.6 × 2.8 µm ; Qe = 11.3
תמונות/קולות

מתצפת.ת
peptolabתיאור
Spirostomum teres Claparède & Lachmann, 1859 from Pussy's Pond, a brackish offshoot of the estuary Acabonac Harbor. Imaged in Nomarski DIC on Olympus BH2 using SPlanapo 20 0.75, SPlanapo 40 0.95 objectives and Splan 100 1.25 oil objective plus variable phone camera cropping on Samsung Galaxy S9+. The cells measure from 480 up to 560 um in average length, have a compact ellipsoid macronuclear nodule, and are brownish in color and highly spirally contractile. A single row of well-developed oral membranelles defines the left side of the long, thin peristomial field, which runs parallel to the main body axis from the anterior end to the cytostome, which is located at 45% of the body length. One stripe of packed cortical granule rows (average 3) between each kinety pair; cortical granules are of different sizes. The contractile vacuole is single and posteriorly located, with a long collecting canal extending along the dorsal side often reaching the anterior end.
"Spirostomum is a genus of ciliated protists that belongs to the class Heterotrichea. It is known for being very contractile. Having been first identified by Christian Gottfried Ehrenberg in 1834, further research has identified eight additional true morphospecies. This bacterivore genus mainly lives in the sediment deposits at the bottom of various aquatic habitats, and members possess rquA genes that could be responsible for their ability to survive in these hypoxic and anoxic environments. They are identifiable by their relatively (to other ciliates) large tubular/flat vermiform bodies. Their life cycle consists of a growth stage, in which they mature, and asexual and sexual reproduction stages. Some species are model organisms for studies on human pathogenic bacteria, while others are sensitive and accurate bioindicators for toxic substances" (1).
"Spirostomum Ehrenberg, 1838 are conspicuous ciliates protists that are easily recognized by their large sizes (500-1000 µm) and elongate bodies, being easily confounded with small helminths. The name Spirostomum refers to the ability these ciliates have to contract in a spiral mode. This type of contraction is due to the presence of post-ciliary, sub-pellicular fibers that arise on the anterior end and spiral in a counterclockwise direction toward the posterior end of the body. Of the eight species that currently comprise Spirostomum, five (including S. minus) possess a moniliform macronucleus" (2). "There are currently eight accepted morphospecies in the genus: S. ambiguum, S. minus, S. teres, S. yagiui, S. dharwarensis, S. semivirescens, S. subtilis, S. caudatum" (1). Among all species of Spirostomum, only S. teres and S. caudatum have a compact macronucleus. S. caudatum is easily separated from S. caudatum by the latter's long thin tail (1). Another old species similar to S. teres is S. ephrussi but according to Repack and Isquith (1974), S. ephrussi is junior synonym of S. teres" (2).
"Spirostomum teres Cláparéde and Lachmann, 1858-1859. [syn: S. ephrussi Delphy, 1939] 150-650 um (avg. 250-450) um long. Length: width ratio about 5-16 (avg. 8-12). 5-15 (avg. 7-10) kineties on each side; usually homogeneous cortical granule rows, variable in number per stripe (2-4). Peristome from 1/3 to slightly more than 1/2 of the body length. Contractile vacuole usually less than 1/5 of the body length. Often brownish. Ellipsoid macronucleus (length: width ratio < 5) in the middle sector of the body, about 20-50 x 5-20 um when stained by Feulgen reaction. A few (1-3) micronuclei 1-2 m long. Molecular analyses suggest that this morphospecies include phylogenetically diverse lineages, some of which are more closely related to S. yagiui and S. dharwarensis; no reliable morphological autapomorphy has yet been detected for these lineages. Found in both fresh- and brackish-water environments. Reported in Europe, central Africa, Madagascar, USA, Brazil, Caspian Sea, India, China and Korea. It sometimes harbors cytoplasmic prokaryotic symbionts" (3).
"Differential diagnosis: 1) Size in vivo 150-600 x20-75 um, mostly 200-400 um, according to DRAGESCO (1960) brackish water forms even reach a length of 880 um. Very contractile; contractile cells more clearly spindle-shaped. 2) Shape moderately broadly cylindrical, somewhat expanded in the middle third, 10-15 times longer than wide. Anterior half distinct, posterior slightly narrowed. Front end slightly beaked, rear end broadly truncated. 3) Macronucleus ellipsoid, about 25-48 x 9-12 um in size, lies in the middle third. 1-2 lenticular, 2-4 um large micronuclei. 4) Contractile vacuole at the posterior end, with a long collecting duct extending forward along the dorsal side. 5) Close under the pellicle, very many spherical, arranged in elongated bands of 2-3 rows each lemon-yellow granules (oil immersion), which give the cell a yellowish tinge. 6) About 25-30 rows of cilia, few in stretched cells, strong in contracted individuals run spirally. 7) The adoral membranelle zone extends from the anterior end to about the middle of the body and bends to the right at the lower end. Oral groove bordered on the right by an undulating membrane. 8] Movement slipping, wriggling and crawling. Floats with rotation around the longitudinal axis" (4).
1, https://en.wikipedia.org/wiki/Spirostomum#cite_note-:3-1
-
Morphology and 18S rDNA gene sequence of Spirostomum minus and Spirostomum teres (Ciliophora: Heterotrichea) from Rio de Janeiro, Brazil. Noemi M. Fernandes & Inácio. D. da Silva Neto. ZOOLOGIA 30 (1): 72–79, February, 2013 http://dx.doi.org/10.1590/S1984-46702013000100009
תמונות/קולות

מתצפת.ת
mnold1תיאור
Mag. 400x
- A water sample taken on 03/31/2024, from a small vernal stream in a wooded area, using a small sample bottled attached to an extension pole. Air temp 46F.
I'm not certain about the ID, but will check with geraldojpr and roman_romanov. Most fascinating about this specimen is that one semi-cell is 3-radiate (3 lobes) and the other is 6-radiate. An intermediate polymorph! Will its progeny be 3- or 6 radiate or a hybrid of both? Interesting! S. margaritaceum is noted to be "variable in the in the number of radial processes, , with 4-6 being the most often found" (https://www.outerhebridesalgae.uk/desmids/desmid-species.php?id=359) ... I could not find a reference noting 3-radiate S. margaritaceum. The first composite image shows the specimen in lateral view, then beginning to roll backwards. The second composite of 9 photos shows the same specimen making a ~180⁰ rotation about a slanted axis. (Panels 8 and 9 vary only in depth of field, not position of the specimen.) Too cool!
For more reference images of S. margaritaceum see:

תמונות/קולות

מתצפת.ת
mnold1תיאור
Mag. 200x
- A water sample taken on 03/31/2024, from a small vernal stream in a wooded area, using a small sample bottled attached to an extension pole. Air temp 46F.
The long neck and specialized oral apparatus suggest membership in the genus Lacrymaria. (Not Lacrymaria olor since it lacks an oral dome and no evidence longitudinal pellicle strips.?) - NOT Lacrymaria! See diagnosis by Bruce Taylor below - Two specimens were observed in the same field of view, one 320µ (partially extended) and a much smaller specimen at 150µ (also partially extended). As they neared each other (no contact occurred) a wonderful little pas de deux ensued. Perhaps depth of field buffered their encounter more than I realized (one individual is usually in the background and out of focus, difficult to discern... but it's there!). After a short dance of to-and-fros and dips of the head, they parted ways seemingly without ever acknowledging the presence of the other. In this photo series, documentation of the pas de deux begins at image 7.
תמונות/קולות

מתצפת.ת
ikhomתיאור
On dead, corticated Jack Pine branches, tiny, black cup fungi. Apothecia sessile.
Asci with many globose spores, IKI+.
Ascospores about 2um in diameter.


תמונות/קולות
מתצפת.ת
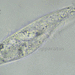
closterium_mysteriumתיאור
Video: https://youtu.be/OYakkMPRt60
Sampling location: The soil sample was collected from the bank of a pond on the Bagaryak River, near the water. The soil was wet but gradually dried out.
Date and time of collection: September 8, 2023
The sample was stored in a sealed glass container at room temperature.
On November 20, 2023, at 1 PM, a small amount of this soil was mixed with clean water.
Date and time of observation: November 25, 2023, at 8 AM
תמונות/קולות

מתצפת.ת
mnold1תיאור
Mag. 100x (3), 400x (1,2,4,5)
Large ciliate (though the anterior portion looks worm-like). Three specimens: A in photos 1&2, B and C in photos 3,4, & 5. All 3 specimens traveled exclusively in a semi-transparent, mucilaginous tube; this is especially apparent in the low-power photo, 3.
- After gently pushing aside the cover of lily pads, a pond side water sample was taken on 06/30/2022 using a 10µ dip net to enrich for microbes. Air temp. 81°F.








תמונות/קולות
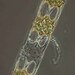
מתצפת.ת
lamawebberתיאור
Species identified: Hobaniella longicrucris P.A.Sims & D.M.Williams, 2018 (previous name is O. longicrucris) (a monospecific genus)
Genus: Hobaniella P.A.Sims & D.M.Williams, 2018
Taxonomic notes on the genera: Hobaniella
Phylum: Bacillariophyta
Class: Bacillariophyta classisincertae sedis
Order: Bacillariophyta ordoincertae sedis
Family: Bacillariophyta familiaincertae sedis
Genus: Hobaniella (Guiry and Guiry 2024)
Type species, synonym(s), etc.: Holotype species: Hobaniella longicrucris (Greville) P.A.Sims & D.M.Williams
Genus summary:
Similar-looking diatoms of the Eupodiscaceae (e.g. Hobanellia, Odontella, Trieres, Zygoceros) and Biddulphiaceae (e.g. Biddulphia) can be approximately divided, morphologically and genetically, into two broad classifications: 1) ocellate and pseudocellate diatoms, respectively. The ocellus-bearing taxa (Eupodiscaceae) are monophyletic, and thus the ocellus is a useful morphological character in establishing the order Eupodiscaceae. However, the Biddulphiaceae are polyphyletic and dispersed across various lineages of multipolar non-pennate diatoms with a taxonomically confused history (Hoban 1983, Round et al. 1990, Ashworth et al. 2013). Previously, Biddulphioid diatoms included most of the Odontella species (Cupp 1943, Shim 1976, Tynni, R. 1986, Waters et al. 1992), including the taxa found in the Salish Sea; there still exists confusion regarding the naming of these two genera, partly because of the morphological variability of species such as O. aurita (Cupp 1943: 616-163). However, in the last twenty years, with molecular and SEM investigations, the taxonomy of ocelli- and pseudocelli-bearing diatoms and the taxonomy of Hobanellia and Odontella have become less ambiguous (Lavigne et al. 2015; Sims et al. 2018; Ashworth et al. 2013; Guiry and Guiry 2024).
Species:
Hobaniella longicrucris P.A.Sims & D.M.Williams, 2018 (basionym: Biddulphia longicruris Greville 1859: 163, pl. VIII [8]: fig. 10; previous name is O. longicrucris)
Description:
Valves broad elliptical-lanceolate. Length of apical axis 15–110μ. The valve face is convex with a prominent domed valve centre positioned between vertical long, narrow elevations with ocellus on each tip. Valve mantle steep, and at the base merging into an expanded hyaline valve margin. External rimoportulae tubular processes 1-3, situated at the apex of the domed valve center, usually long and slender, with rounded fork-like ends and slightly inflated at the base. Valve areolae radiating from a small hyaline central area, forming concentric ellipses on each half of the valve, in nearly parallel rows near base of valve mantle. Areolae on valve 12–17 in 10μ. Areolae occluded by a simple velum. Girdle zone with straight sides, with fine, parallel, vertical rows of pores 18–21 in 10μ. Girdle bands sometimes present, finely poroid 15–18 in 10μ, in vertical rows. Chloroplasts small, numerous, near the wall. Nucleus central. Cells joined into chains by ends of ocellus on valve elevations (Greville 1859: 163; Cupp 1943; Hasle and Syvertsen 1996; Sims et al. 2018. Guiry and Guiry 2024)
Salish Sea specimens: Hobaniella longicrucris-SHW-I-1036-2-July 20-2005_4.tif; Hobaniella longicrucris-SHW-I-1039-2-400x Dialux-July 20-2005_3.tif; Hobaniella longicrucris-SHW-I-s4700-July 14-2005_m007_3.tif; Hobaniella longicrucris-SHW-I-s4700-July 14-2005_m010_4.tif; Hobaniella longicrucris-SHW-Nov 10-2010_0014_4 copy.tif; Hobaniella longicrucris-SHW-Nov 10-2010_0003_4b.tif; Hobaniella longicrucris-SHW-I-s4700-July 14-2005_m004_4.tif.
Morphological data:
Valves broad elliptical-lanceolate. Length of apical axis 29.6-49.1 μm. The valve face is convex with a prominent domed valve centre positioned between vertical long, narrow elevations with ocellus on each tip. Valve mantle steep, and at the base merging into an expanded hyaline valve margin. External rimoportulae tubular processes 2, situated at the hyaline apex of the domed valve center, long and slender, with swollen fork-like tips and slightly inflated at the base. Valve areolae radiating from a small hyaline central area, forming concentric ellipses on each half of the valve, in nearly parallel rows near base of valve mantle. Areolae on valve 12 in 10μ. Girdle zone with straight sides, with fine, parallel, vertical rows of poroids 36 in 10μ. Girdle bands sometimes present, finely poroid, in vertical rows. Chloroplasts small, numerous, near the wall. Cells joined into chains by ends of ocellus on valve elevations.
Odontella_longicruris-SHW-2022, #161 RBCL 283, etc.: (100% 3e and 1e expect values, excellent distance tree results).
Odontella is cosmopolitan in the marine littoral, planktonic, epiphytic and benthic habitats. Odontella aurita (McIntire and Overton 1971, Tynni 1986, Sancetta and Calvert 1988); Hobaniella longicruris (Waters et al. 1992, Pienitz et al. 2003) and O. obtusa (Rao and Levin 1976, Shim 1976) are commonly found species within the Salish Sea and along the west coast of North America (Cupp 1943). These three species are frequently found in the plankton at Spanish Hills Wharf (SHW), Trincomali Channel, Galiano Isl. and occasionally in amongst epiphytes in the marine eelgrass Zostera marina at Montague Harbour Marine Provincial Park (MHMPP), Galiano Isl., BC, Canada.
From a plankton tow at Spanish Hills Wharf (SHW), Trincomali Channel, Galiano Island, BC, Canada, on July 14, 20, 2005. Live imaging with a Leitz Dialux and Nikon CoolPix 4500 camera. SEM imaging with a Hitachi s4700 at the BioImaging Facility, University of British Columbia. Thank Elaine Humphrey for SEM support.
References:
Ashworth, M. P., Nakov, T., & Theriot, E. C. (2013). Revisiting Ross and Sims (1971): toward a molecular phylogeny of the Biddulphiaceae and Eupodiscaceae (Bacillariophyceae). Journal of Phycology, 49(6), 1207–1222. https://doi.org/10.1111/JPY.12131
Guiry, M.D. in Guiry, M.D. & Guiry, G.M. 2021. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 09 October, 2021.
Hasle, G.R. & Syvertsen, E.E. (1996). Marine diatoms. In: Identifying Marine Phytoplankton. (Tomas, C.R. Eds), pp. 5-385. San Diego: Academic Press.
Hoban, M.A. (1983). Biddulphioid diatoms II: The morphology and systematics of the Pseudocellate species, Biddulphia biddulphiana (Smith) Boyer, B. alternans (Bailey) Van Heurck, and Trigonium arcticum (Brightwell) Cleve. Botanica Marina, 26(6): 271-284
Hoppenrath, M., Elbrachter, M., Drebes, G. (2009) Marine Phytoplankton, Selected microphytoplankton species from the North Sea around Helgoland and Sylt. E. Schweizerbart’sche Verlagsbunchhandlung, Stuttgart, Germany.
Lavigne, A.S., Sunesen, I. & Sar, E.A. (2015). Morphological, taxonomic and nomenclatural analysis of species of Odontella, Trieres and Zygoceros (Triceratiaceae, Bacillariophyta) from Anegada Bay (Province of Buenos Aires, Argentina). Diatom Research 30(4): 307-331.
Pienitz, R., Fedje, D. and Poulin, M. (2003) Marine and Non-Marine Diatoms from the Haida Gwaii Archipelago and Surrounding Coasts, Northeastern Pacific, Canada In Bibliotheca Diatomologica (H. Lange-Bertalot and P. Kociolek, eds.), Band 48, J. Cramer, Stuttgart, 146 pp.
McIntire, C. D. and Overton, W. S. (1971). Distributional Patterns in Assemblages of Attached Diatoms from Yaquina Estuary, Oregon. Ecology, Vol. 52, No. 5. pp. 758-777.
Rao, V.N.R. and Levin, J. 1976. Benthic marine diatom flora of False Bay, San Juan Island, Washington. Syesis, 9:173–213.
Round, F.E., Crawford, R.M. and Mann, D.G. (1990), The Diatoms, Biology & Morphology of the Genera, pp. 220-221. Cambridge University Press, Cambridge, UK.
Sancetta, C. and Calvert, S. E. (1988). The annual cycle of sedimentation in Saanich inlet, British Columbia: implications for the interpretation of diatom fossil assemblages. Deep Sea Research Part A. Oceanographic Research Papers, 35(1), 71–90. doi:10.1016/0198-0149(88)90058-1
Shim, J. H. (1976). Distribution and Taxonomy of Planktonic Marine Diatoms in the Strait of Georgia, B.C. Phd. Thesis, UBC.
Sims, P.A. (ed.) (1996). An atlas of British diatoms arranged by B. Hartley based on illustrations by H.G. Barber and J.R. Carter. pp. 406-409, Bristol: Biopress Ltd.
Sims, P. A., Williams, D. and Ashworth, M. P. (2018). Examination of type specimens for the genera Odontella and Zygoceros (Bacillariophyceae) with evidence for the new family Odontellaceae and a description of three new genera. Phytotaxa 382(1):1. DOI: 10.11646/phytotaxa.382.1.1
Spaulding, S.A., Bishop, I.W., Edlund, M.B., Lee, S., Furey, P., Jovanovska, E. and Potapova, M. Diatoms of North America. Retrieved November 6, 2021, from https://diatoms.org
Tynni, R. (1986). Observations of diatoms on the coast of the State of Washington. Geological Survey of Finland. Report of Investigation 75
Waters, R. E., Brown, L.N., and MG Robinson, M.G. (1992). Phytoplankton of Esquimalt Lagoon, British Columbia: comparison with west Vancouver Island coastal and offshore waters. Canadian Technical Report of Hydrography Ocean Sciences 137.
תמונות/קולות

מתצפת.ת
peptolabתיאור
Trachelius ovum EHRENBERG, 1831 EHRENBERG, 1838 from the acidic freshwater kettle pond Chatfield's Hole. Imaged in Nomarski DIC on Olympus BH2S using SPlanapo 20 0.75 and Splanapo 40 0.95 objectives plus variable phone camera cropping on Samsung Galaxy S9+. The cells measure 240 um in length. The following discussions and several figures are adapted from Foissner et al (1995) (1).
According to EHRENBERG (1838), FOISSNER & FOISSNER (1988b) and KAHL (1931a), Trachelius ovum has several safe synonyms, which do not appear in the saprobiological literature: T. cicer SCHRANK- this older synonym was never used and should therefore be suppressed forever for reasons of stability, T. vorax EHRENBERG, Amphileptus rotundus MASKELL, Trachelius Leidyi FOULKE. The exact organization of this common ciliate was previously insufficiently known, although it was reported several times and has been processed using modern methods (DRAGESCO & DRAGESCO-KERNEIS 1986, FRYD VERSAVEL et al. 1975, SONG & WILBERT 1989). The representations are too schematized and incorrect in detail. We have therefore reworked T. ovum for the “Atlas”. This showed that the somatic and oral ciliation is very similar to that of Dileptus. Nevertheless, Trachelius is clearly demarcated from Dileptus namely by the ventro-lateral fossa, where the ciliation is slightly modified and the club-shaped mouth funnel, which consists of a thick layer of the finest fibrils.
Congener comparison: Trachelius subtilis PENARD, which has not yet been sufficiently confirmed, has only 12 contractile vacuoles and no suction cup. Dileptus species usually have a clearly pointed rear end and are always much slimmer. Paradileptus elephantinus lives predominantly in the pelagic of stagnant waters and has a rosary-shaped (moniliform) macronucleus. The characteristics are particularly important for identification are 2, 3, 4 .
Interestingly, I found one individual with two closely abutting oval macronuclei (see end of pictures after sampling site pictures). Trachelius subtilis PENARD, 1922, a species which Vďačný & Foissner (2) synonymized with T. ovum, is stated to have two oval macronuclear nodules. However, Martin Kreutz disagrees with Vďačný & Foissner and writes: "Trachelius subtilis was first described by Penard (1922). The species is smaller than Trachelius ovum and its main characteristic is a two-part macronucleus with a spherical micronucleus in the middle. In 2012 Vďačný & Foissner (2) synonymized Trachelius subtilis with Trachelius ovum with the argument that Penard (1922) possibly only found specimens of Trachelius ovum with a macronucleus constricted in the middle and that this is an observational error. Vďačný & Foissner obviously did not find any specimens with such a constricted macronucleus themselves. However, Penard was a very precise observer and Kahl (1935) also found specimens of Trachelius subtilis, which confirmed and supplemented Penard’s results" (3). Dr. Kreutz's examples do show two clearly separate macronuclear nodules with an intervening micronucleus in contrast to my observation of two closely abutting macronucleuar nodules with no visible micronucleus so I cannot rule out a folded dumbbell-shaped macronucleus appearing as two nodules in my example.
Differential diagnosis
Size in vivo 200-600 x 75-350 um, usually 250-350 um long.
Shape sac-shaped to almost spherical, starving specimens clearly flattened on one side. Proboscis often only about 1/4-, rarely up to 1/2-length, usually curved dorsally. In well fed specimens it becomes a short, stalk-shaped extension. Ventro-laterally a small, difficult to recognize pit that serves as a suction cup.
Macronucleus dumbbell-shaped, often disintegrates into a few spherical parts in postconjugates. Several micronuclei.
Many small contractile vacuoles scattered throughout the cell. Plasma very strongly vacuolated, the strands form a coarse network.
Short, rod-shaped extrusomes in the proboscis along the ridge of the mouth. Cortex thick, with many ellipsoid granules.
About 80-120 longitudinal rows of cilia, some of which extend into the ventro-lateral fossa, where there are several specializations, which are explained in the figure legends. Brush 3-4 rows, on the dorsal side of the trunk, a row extending almost to the end of the body; only clearly visible after silver impregnation and in the scanning electron microscope.
Mouth entrance at the base of the proboscis, surrounded by many very delicate bars that form a club-shaped, thick-walled funnel. To the right of the circumoral row of eyelashes there is a longitudinal row of cilia, on the left there are many short oblique rows of perioral cilia; more precise structure of the oral cilia can only be recognized after silver impregnation.
- FOISSNER W., BERGER H., BLATTERER H. & KOHMANN F. (1995): Taxonomische und ökologische Revision der Ciliaten des Saprobiensystems – Band IV: Gymnostomatea, Loxodes, Suctoria. – Informationsberichte des Bayer. Landesamtes für Wasserwirtschaft, 1: pp. 208-18
- VĎAČNÝ P. & FOISSNER W. (2012): Monograph of the dileptids (Protista, Ciliophora, Rhynchostomatia). – Denisia, 31: 1–529.
- https://realmicrolife.com/trachelius-subtilis/

תמונות/קולות
מתצפת.ת
connorquinnתיאור
Conidium stained with lactophenol/cotton blue. Retrieved from foam sample.
תמונות/קולות
מתצפת.ת
peptolabתיאור
Remanella granulosa (Kahl, 1933) Foissner, 1996 from the intertidal benthos of a small estaurine beach near the boat basin at Moneybogue Bay in Westhampton Beach on the south shore of Long Island. It is imaged in Nomarski DIC using Olympus BH2S under SPlanapo 40 0.95 and SPlanapo 20 0.75 objectives plus variable phone cropping on Samsung Galaxy S9+. The cells range from 180 up to 400 um in length, contain an average of 12 up to 14 Müller vesicles in varying stages of development with 5 Müller vesicles in a row at the head end, and three macronuclei with a single micronucleus. These features point to a diagnosis of R. granulosa, which is the only species of this size listed in the review of Xu et al (2012) (1) to have three macronuclei. Their population had individuals possessing up to 14 Müller vesicles which I also found fairly consistently in my population with the more posterior Müller vesicles being less developed. Ma et al (2022) (2) described a new species, Remanella elongata, with similar size and also having similar range of Müller vesicles (7-13) and from 2 to 3 macronuclei, but this species has a prominent longitudinal glabrous stripe lacking in my population of R. granulosa.
Improved diagnosis by Xu et al (2015) "Cell size in vivo about 150–400 × 25–45 µm; ratio of buccal field/body length about 1/4–1/5; 19–23 right lateral ciliary rows; right buccal, left outer buccal and intrabuccal kineties composed of 140–189, 35–70 and 64–78 dikinetids respectively; two or three macronuclei and a single micronucleus; cortical granules brown in colour, present on both sides of body and densely packed in buccal area and around pharyngeal tube" (1).
"Body slender and flexible, mostly about 300 × 40 µm in vivo, with buccal field occupying c. 1/4–1/5 of body length. Cilia about 8 µm long. Cells usually appear dark brownish at low magnification due to brown cortical granules which are round, c.0.5µm in diameter, and densely packed in buccal area, around pharyngeal tube, and on both sides of cell. On right side of cell granules seem to be arranged in lines along ciliary rows. Usually eight or nine Müller vesicles located near dorsal margin, each c.8µm in diameter and with a globular or ellipsoidal mineral granule c. 4µm across. Some individuals possessing up to 14 Müller vesicles. Cytoplasmic spicules about 10–12 µm long, scattered throughout cell. Two or three macronuclei with a single micronucleus located between them. Locomotion by gliding between sand grains or along bottom of Petri dish" (1).
"Remanella granulosa was first described by Kahl (1933) as follows: ‘body size 150–300µm; cortical granules brown, packed in buccal area. Considering these characters and the general appearance of the figure given by Kahl (1933), the Chinese population corresponds well to the original report. The main difference between these two populations is the number of Müller vesicles (5 according to the original illustration vs.about 8 or 9 in the Chinese population).Based on our observations, however, the number of Müller vesicles is somewhat variable in this species. Therefore we believe the two populations are conspecific. The Chinese population also matches that described by Dragesco (1960) from general appearance. The ultrastructure of the nuclear apparatus and endoplasm of R. granulosa were reported by Raikov(1994) and Raikov& Kovaleva( 1990)" (1).
- Diversity of the karyorelictid ciliates: Remanella (Protozoa, Ciliophora, Karyorelictida) inhabiting intertidal areas of Qingdao, China, with descriptions of three species. XU ANXU,MIAO MIAO,ALAN WARREN & WEIBO SONG. Systematics and Biodiversity (2012), 10(2): 207–219
- Molecular phylogeny and taxonomy of four Remanella species (Protozoa, Ciliophora): A flagship genus of karyorelictean ciliates, with descriptions of two new species. Ming-Zhen Ma, Yu-Jie Liu, Yuan Xu, Bo-Rong Lu, Yu-Qing Li, Saleh A. AL-Farraj, Giulio Petroni, Wei-Bo Song, Ying Yan. Zool. Res. 2022, 43(5): 827−842
תמונות/קולות

מתצפת.ת
samuelbrinkerתיאור
Perithecia with furrowed neck. Spores muriform, 2/ascus, 49-58 x 20-23 um.

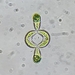
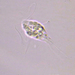
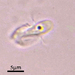
תמונות/קולות
מתצפת.ת
closterium_mysteriumתיאור
Video: https://youtu.be/G4yMvNvUhdw
Sampling location: A water sample was collected from the bank of the Ufa River within the boundaries of Nyazepetrovsk town.
Date and time of collection: 18 Jun 2023 at 6 PM
Date and time of observation: 25 Jun 2023 at 5 PM
The sample was stored at room temperature in a plastic container.
תמונות/קולות
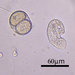
מה
סוג Colpodaמתצפת.ת
crseaquistתיאור
Gathered dry leaves on 2024-02-23 and stored in water.
Two daughters emerging from a division cyst:
- Initial observation of cyst with another individual
- 2 minutes later
- 18 minutes later
- 77 minutes later
- 84 minutes later
- 140 minutes later
- 153 minutes later
- 154 minutes later 1st daughter emerges
- 155 minutes later
- 162 minutes later
- 163 minutes later 2nd daughter emerges
- 2nd daughter swims away
Video: https://youtu.be/vntikKDmSDE

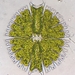
תמונות/קולות

מתצפת.ת
lamawebberתיאור
Species identified:
Shionodiscus poro-irregulatus (Hasle & Heimdal 1970) Alverson, (Kang et Theriot (2006)
Genus:
Taxonomic classification:
Phylum: Heterokontophyta
Subphylum: Bacillariophytina
Class: Medioiophyceae
Subclass: Thalassiosirophycidae
Order: Thalassiosirales
Family: Thalassiosirosiraceae
Genus: Shionodiscus (Guiry and Guiry 2024)
Type species: Shionodiscus oestrupii (Ostenfeld) A.J.Alverson, S.-H.Kang & E.C.Theriot 2006: 258 (Guiry and Guiry 2024)
Genus Summary:
A small marine genus similar in appearance to Thalassiosira Cleve. However, it differs from the later genus by the labiate process always being some distance from the margin, normally on the valve face, occasionally near the edge of the face. The strutted processes are typically long inward extensions and always the outward extensions are reduced or absent (Hasle and Syvertsen
1996; Alverson et al. 2006; Wilks and Armand 2017).
Species:
Shionodiscus poro-irregulatus (Hasle & Heimdal 1970)
Alverson, (Kang et Theriot (2006)
Thalassiosira proro-irregulata Hasle & Heimdal 1970
(Hasle & Heimdal, 1970, p. 573-574, Fig. 55-64, 71, 72.)
Description:
Girdle view: Diameter 21-30 µm. Valves flat, slightly depressed at the center, with slightly rounded edges. Cells close together in chains. Distances between cells about one-fifth of the pervalvar axis and less than one-fourth of the diameter. Chloroplasts numerous. Valve view: Valve areolae is 10 - 15 in 10 µm. Hexagonal areolae, approximately same size throughout the valve. Inner sieve membrane of areolae has numerous small pores close together. 1-8 central trifultate strutted processes, independent of cell size, irregularly scattered in the valve centre. Marginal processes, internally long, not extending on exterior of the frustule, 4-6 in 10 µm. Marginal processes situated vertically on a high and steep mantle. Labiate process located mid-way between margin and center. Labiate process is notably anvil shaped. Labiate process is 5-6 areolae from central process. (Hasle and Heimdal 1970; Hasle and Syvertsen, 1996; Kang et Theriot 2006; Guiry and Guiry 2024)
Salish Sea specimens: Shionodiscus poro-irregulatus Gen Plank (THAL)-Box 19 June 25-2022-TM4000_nitric-Aug 14-2022_ Stb Site 1b_60(x3k)4.tif; Shionodiscus poro-irregulatus-Gen Plank-Porlier-(THAL)-Box 19 June 25-2022-TM4000_nitric-Aug 14-2022 Stb Site 1b_57(x1.8k)_4.tif.
Morphometric data:
Girdle view: Diameter 21.9-37.5 µm. Valves flat, slightly depressed at the center, with slightly rounded edges. Valve view: Valve areolae is 8-12 in 10 µm. Hexagonal areolae, approximately same size throughout the valve. 2-5 central trifultate strutted processes irregularly scattered in the valve centre. Marginal processes, internally long or short, but not extending on exterior of the frustule, 4-5 in 10 µm. Marginal processes situated vertically on a high and steep mantle. Marginal processes are 1.8-2.4 µm apart. Labiate process located 50-60% towards centre of valve. Labiate process is notably anvil shaped. Labiate process is 5-6 areolae from central process.
This is a first report of Shionodiscus poro-irregulatus in the Salish Sea and Pacific Western Canada.
From a general 60 µm mesh plankton tow at Porlier Pass, Galiano Island, BC, Canada, on June 25, 2022. After 4% formalin (final concentration) fixation, cells were cleaned with concentrated nitric acid on 12 mm glass coverslips and rinsed in distilled water deionized water. Mounted with double sided taper to 13 mm aluminum SEM stubs. Thank you to Andrew Simon of IMERSS for collecting the sample. SEM imaging with a Hitachi TM4000 at the Advanced Microscope Facility, UVIC. Thank you Hitachi HiTech and Elaine Humphrey for SEM support. Images by Arjan an Asselt, Melanie Quenneville and Mark Webber. Specimen preparation, imaging and taxonomy by Mark Webber (imerss.org).
References:
Alverson, A.J., Kang, S.-H. & Theriot, E.C. (2006). Cell wall morphology and systematic importance of Thalassiosira ritscheri (Hustedt) Hasle, with a description of Shionodiscus gen. nov. Diatom Research 21(2): 215-262.
Fryxell, G. A., & Hasle, G. R. 1979. The genus Thalassiosira: T. trifulta sp. nova and other species with tricolumnar supports on strutted processes. Beiheft zur Nova Hedwigia 64:13-32.
Fryxell, G. A., & Hasle, G. R. (1980). The marine diatom Thalassiosira oestrupii" Structure, taxonomy and distribution. American Journal of Botany 67:804-814.
Hasle, G.R. & Syvertsen, E.E. (1996). Marine Diatoms. In: Identifying Marine Phytoplankton. (Tomas, C.R. Eds). San Diego: Academic Press.
Guiry, M.D. & Guiry, G.M. 2007, AlgaeBase version 4.2. World-wide electronic publication, National University of Ireland, http://algaebase.org, searched March 15, 2024.
Li, Y. and Lu, S. (2013). The genus Thalassiosira off the Guangdong coast, South China Sea. Botanica Marina; 56(1): 83–110. DOI: 10.1515/bot-2011-0045
Wilks, J. V. and Armand, L. K. (2017): Diversity and taxonomic identification of Shionodiscus spp. in the Australian sector of the Subantarctic Zone, Diatom Research, DOI: 10.1080/0269249X.2017.1365015


תמונות/קולות

מתצפת.ת
mossgeekתיאור
First occurance in southern California since 1885. J-Dar also found one in Santa Barbara area.
תמונות/קולות

מתצפת.ת
lamawebberתיאור
Species Identified:
Hemiaulus sinensis (chinensis) Greville 1865
Genus: Hemiaulus Heiberg, 1863, nom. cons. (Guiry and Guiry, 2020)
Heiberg, P.A.C. (1863). Conspectus criticus diatomacearum danicarum. Kritisk oversigt over de danske Diatomeer. pp. 1-135, 6 pls. Kjøbenhavn: Wilhelm Priors Forlag.
Taxonomic notes on the genus Hemiaulus:
Phylum: Bacillariophyta
Subphylum: Bacillariophytina
Class: Mediophyceae
Subclass: Chaetocerotophycidae
Order: Hemiaulales
Family: Hemiaulaceae
Genus: Hemiaulus (Guiry and Guiry, 2024)
Type species, synonyms, etc.: Hemiaulus proteus Heiberg 1963: 47; pl. I, fig. I
Genus Summary:
Hemiaulus is a cosmopolitan marine planktonic genus found frequently in the fossil record. United by short or long processes, forming straight or curved chains, and occasionally rotated about the long axis. Cells are lightly silicified. Valves are elliptical and have two long thin processes ending in apical spines with claw like tips. The tips are often flattened with pointed ends that link adjacent cells without pseudocelli or swollen ends as found in Eucampia. The valve face is curved and merges gradually into the deep mantles. The areolae are simple round pores (H. hauckii) or large elliptical to rectangular holes closed by complex cribra. The single rimoportula is often present in a central or offset position or occasionally absent. Girdle bands (copulae) split with pointed ends and finer areolate than on the valves. Plastids discoid and small
(Cupp 1943, Ross, Sims and Hasle, 1977; Ross and Sims, 1985; Round et al. 1990;. Hasle and Syvertsen 1996).
Species Identified:
Hemiaulus sinensis (chinensis) Greville 1865: 5, pl. V [5]: fig. 9
Description:
In valve view the cells are broadly elliptical in outlines with a slightly convex surface. The apical axis is 15–36 μm long. Chains are straight or curved. Pervalvar axis generally elongated. The valve mantle is high. The two external processes on the valves are moderately thin, somewhat truncated but robust, with strong claws-like structures on the ends. Compared with H. hauckii, the frustule is more siliceous. The areolae are round in a radial pattern and on the mantle surface the rows are in a distinctive eccentric arrangement; the center of the areolation does not coincide with the center of the valve but lies on one of the mantle surfaces. Areolae in the center of the valve 7–9 in 10 μm, on the base of the mantle 11–13 in 10μm. Very fine punctation present on the bands (copulae) 28–29 striae in 10μm (Greville, 1865: 5; Pl. 5, fig. 9; Cupp 1943; Hasle and Syvertsen 1996; Al-Kandari 2009).
Morphometric description of Trincomali Channel specimen: Hemiaulus sinensis-isolate-SHW-Jan 14-2024 (Feb 7-2024) antibiotic_AE31_0002_2.tif; Hemiaulus cf. sinensis-SHW-Culture - Feb 29-2024-E800_100x oil_AvA_0051_4 copy.jpg.
Morphometric data:
In valve view the cells are broadly elliptical to rectangular in outline with a slightly convex surface. Apical axis is 26 μm long. Chains are curved. Pervalvar axis generally elongated. The valve mantle is high. The two external processes on the valves are moderately thin, somewhat truncated but robust, with strong claws-like structures on the ends. Areolae on the base of the mantle 14 in 10μm
Reported by Shim (1976) in the Strait of Georgia, Salish Sea.
Methods:
General plankton sample with a 20 µm net at Spanish Hills Wharf, Trincomali Channel, Galiano Island, Gulf Islands, BC, Canada, January 14, 2024. Single isolate grown in f/2 media with LED lights and images of the original colony islolated. Dead cells mounted in sterile marine water between a coverslip and slide and imaged with a Nikon E800, 100x DIC NA 1.40 oil lens and an MU2003MP camera. Collecting, imaging, isolating and cloning by Arjan van Asselt. Taxonomy by Mark Webber.
References:
Al-Kandari, M., Al-Yamani, F.Y. and Al-Rifaie, K. (2009). Marine Phytoplankton Atlas of Kuwait’s Waters. Kuwait Institute for Scientific Research, Kuwait City, 351 p.
Cupp, E. E. (1943). Marine Plankton Diatoms of the West Coast of North America. Bull. Scrips. Inst. Oceanography. 5: 1-238.
Greville, R.K. (1865). Descriptions of new genera and species of Diatoms from Hongkong. Annals and Magazine of Natural History, series 3, 16(91).
M.D. Guiry in Guiry, M.D. & Guiry, G.M. August 25, 2020. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org; searched on Feb. 8, 2024.
Hasle, G.R. & Syvertsen, E.E. (1996). Marine diatoms. In: Identifying Marine Phytoplankton. (Tomas, C.R. Eds. San Diego: Academic Press.
Hendey, N.I. (1964). An introductory account of the smaller algae of British coastal waters. Part V: Bacillariophyceae (diatoms). pp. [i]-xxii, 1-317. London: Ministry of Agriculture, Fisheries and Food, Fishery Investigations. Her Majesty’s Stationery Office.
Mather, L., MacIntosh, K., Kaczmarska, I., Klein, G. & Martin, J.L. (2010). A checklist of diatom species reported (and presumed native) from Canadian coastal waters. Canadian Technical Report of Fisheries and Aquatic Sciences 2881: 1-78.
Ross, R., Sims, P. A., & Hasle, G. R. 1977. Observations on some species of the Hemiauloideae.
Beiheft zur Nova Hedwigia 54:179-213.
Ross, R. & Sims, P.A. (1985). Some genera of the Biddulphiaceae (diatoms) with interlocking linking spines. Bulletin of the British Museum (Natural History) Botany Series, 13(3): 227-381, 33 pls.
Round, F.E., Crawford, R.M. & Mann, D.G. (1990), The Diatoms, Biology & Morphology of the Genera, pp. 580-581. Cambridge University Press, Cambridge, UK.
Shim, J. H. (1976). Distribution and Taxonomy of Planktonic Marine Diatoms in the Strait of Georgia, B.C. Phd. Thesis, UBC.

תמונות/קולות

מתצפת.ת
closterium_mysteriumתיאור
Video: https://youtu.be/1Q8kDGQLsMs
The pseudoscorpion was discovered on a windowsill adjacent to a container of forest soil, vegetation, and springtails. Therefore, it is unclear whether the pseudoscorpion was an inhabitant of the apartment or brought in from the forest.



